Quantitative phase imaging is a powerful, non-invasive and quantitative microscopy technique that allows multi-scale imaging of cellular systems, explain Mustafa Mir and Gabriel Popescu

The development of light microscopy in the 17th century led directly to the field of cell biology and transformed our view of life. It was quickly realized, however, that biological samples are optically transparent and therefore difficult to image using visible light. Indeed, in 1873 Ernst Abbe showed that there was a fundamental diffraction limit to optical resolution – a result that would later be more deeply understood in terms of Heisenberg’s uncertainty principle. For most of the 20th century the resolution dogma remained unchallenged and cell microscopy witnessed a constant struggle to improve image resolution.
Contrast, on the other hand, can be improved by using “exogenous” agents – those introduced from outside the body – to label structures of interest. Stains are essential for histopathology, for example, while fluorescent dyes lie behind many recent advances in cell biology. Although these techniques provide high-contrast images, samples tend to suffer from phototoxicity or photobleaching, which can damage the cells under investigation and limit continuous measurement times, respectively. An alternative way to improve contrast is to engineer optical systems that exploit the scattering interaction between light and the sample to reveal structure.
Imaging breakthroughs
The first breakthrough for such label-free, or “endogenous”, contrast imaging came in the 1930s with the invention by Dutch physicist Frits Zernike of phase contrast microscopy (PCM). This allowed fine biological structures to be visualized with sub-cellular resolution. Although PCM enabled biologists to observe living cells for long periods, and earned Zernike the 1953 Nobel Prize for Physics, the phase information is not retrieved quantitatively and therefore provides little more than a high-contrast image. If we are to truly characterize biological systems, we need high-throughput, quantitative methods that perturb samples as little as possible and which can quantify dynamic behaviour at temporal scales ranging from milliseconds to days, and spatial scales ranging from nanometres to centimetres.
A new technology called spatial light interference microscopy (SLIM) incorporates all of these key capabilities. SLIM is a type of quantitative phase imaging (QPI) – a field born out of Dennis Gabor’s pioneering work in holography in the 1940s. It measures phase shifts through the specimen at each point in the field of view to provide unique, quantitative insight into biological systems in a completely non-invasive manner (Optics Express 19 1016). In order to retrieve phase information, two waves are required: one that propagates through the sample and one that does not, after which the two wave fronts can be compared. While such interferometry is already used in cell microscopy, for example via existing PCM technology, these are not quantitative.
In traditional interferometry, the sample and reference fields travel through different optical paths. That makes it hard to extract phase information because noise experienced by the reference field is uncorrelated with that experienced by the sample field. Furthermore, interferometers traditionally use lasers – a highly coherent source of light that creates random interference patterns, known as “speckle”, which makes the technique less spatially sensitive.
SLIM avoids uncorrelated noise by making the reference and sample fields travel through the same path, such that the noise cancels out when they interfere. By using a broadband source with a shorter coherence length, it also addresses the problem of speckle. Taking the form of an add-on module for existing commercial PCM devices, the SLIM system builds on these basic principles of the phase-contrast microscope and transforms it from a qualitative to a quantitative instrument.
Back in the 1930s, Zernike had appreciated that light emerging from the sample is spatially decomposed into two parts. There is a low-frequency portion that passes through the sample undeviated, which falls in the centre of the back focal plane of the objective, and a high-frequency component surrounding the non-deviated light. In the case of thin biological specimens, the phase shift induced by the sample relative to the background barely modifies the image intensity. But Zernike realized that by introducing a thin phase plate to create a 90° phase delay to the background light, minute changes in phase translate to a detectable change in intensity.
SLIM in action
SLIM turns Zernike’s phase plate into a tunable element. It uses a lens to project the “pupil plane” of the microscope onto a spatial light modulator that imparts a phase delay to the incident field. A second lens is then used to project the modulated image onto a CCD camera. Four intensity images are acquired at 90˚ phase-delay increments between the scattered and unscattered fields, allowing the quantitative phase image to be calculated. The apparatus therefore offers all the bells and whistles of the modern microscope – such as automated positioning, stage-top incubation and fluorescence capabilities – while providing unprecedented quantitative insight into cell and tissue physiology.
Since the measured phase shift is proportional to the product of the relative refractive index and the thickness of the sample, SLIM has two natural modes: to measure topography in situations where the refractive index is known, and vice versa. One of the first samples we measured, in 2008, was a carbon film grating with a known refractive index, which proved that the topography measured by SLIM agrees with measurements using atomic force microscopy to within a fraction of a nanometre. The broadband illumination led to a spatial sensitivity of 0.3 nm, while SLIM’s common path geometry enabled a temporal sensitivity of 0.03 nm.
Remarkably, when it comes to living cells, the phase information can also be used to calculate the non-aqueous content, or “dry mass”, at the single-cell and even sub-cellular level – a fact first realized by biologist Robert Barer in the 1950s. Our experiments show that SLIM is sensitive to changes in dry mass at the femtogram level and so can be used to study the growth characteristics of single living cells. Since cells weigh on the order of picograms and only double their mass over the cell cycle before dividing, femtogram accuracy is vital to know if a single cell is growing exponentially or linearly, for instance. Recently, we used SLIM to measure the growth of a single E. coli bacterium and verified that our results agreed well with previous growth measurements from flow cytometers and microresonators (Lab on a Chip 14 646).
Mapping mammalian cell growth
We then turned our attention to the growth of single mammalian cells as a function of cell cycle – a problem that at the time could not be addressed with any other system. Such cells undergo a cell cycle that typically starts off with a growth phase, followed by a DNA replication phase, then a second growth phase, and finally mitosis or cell division. Researchers typically use samples with artificially synchronized population cycles, the effects of which are transient and perturbative. We used SLIM to acquire fluorescent images using a reporter that indicates when cells are in the DNA replication phase, and found that mammalian cell growth is much more variable than the traditional textbook picture (Proc. Natl Acad. Sci. 108 13124).
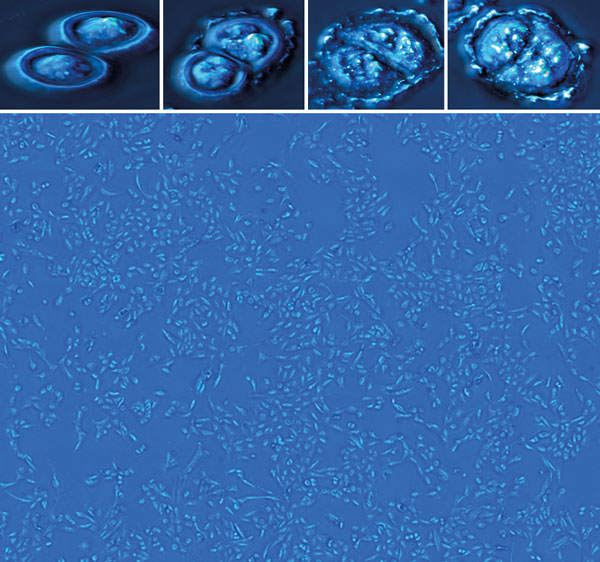
More recently, we have used SLIM to characterize the mass growth during neural‑network formation in mature human neurons over a period of 24 hours. The results revealed a period of growth lasting 10 hours, during which the SLIM images showed neurons extending and exploring their environment. This was followed by a plateau characterized by an aggregation of clusters and finally larger-scale reorganization. We then analysed the data using a new technique called dispersion-relation phase spectroscopy (DPS). This technique analyses fluctuations of mass as a function of time and space, providing a quantitative picture of mass transport in cells (Optics Express 19 20571).
Applying DPS at 0 and 24 hours we found that the mass transport characteristics of the neuron culture shift from being dominantly diffusive to dominantly directed as the network matures and forms more connections. This indicates that the network at some point “decides” to divert energy from growth and exploration to transport and communication (Scientific Reports 4 4434).
The broadband nature of the illumination in SLIM, coupled with a high numerical aperture, also enables high-resolution single-cell tomography by mapping the refractive index in 3D using a method called white-light diffraction tomography. As recently demonstrated by our group, this allows the study of 3D biological structures in living cells – a feat that was previously only achievable using fluorescence-based methods such as confocal or light-sheet microscopy (Nature Photonics 8 256).
Although many biological applications of QPI have already been explored, we believe that the field is just getting started. Recent results on SLIM imaging of biopsies, for instance, indicate that clinical information can be extracted from the nanoscale morphology of tissues (Scientific Reports at press). Until now, SLIM has been used mainly by optical engineers and physicists, so we need to further distill the data into parameters that directly provide insight to biologists and to identify relevant research questions. The recent commercialization of SLIM technology by Phi Optics – a spin-out from the University of Illinois – is a major step forward in this regard, since it adds QPI capabilities to any existing microscope.
As well as getting the instruments into the hands of more biologists, we also need to develop mathematical tools to analyse the wealth of information in QPI data. One such approach aims to connect the large, multi-parameter space measured by QPI to cellular phenotype, which is currently measured using antibody-based labels. If successful, this could ultimately remove the need for fixing and staining, in the process saving researchers thousands of dollars. Given the pace of growth and rapid advancements in the field, we believe that QPI techniques will play a dominant role in the future for many biologists and biophysicists working to elucidate the mysteries of cellular life.